In our last LEMONTREE science meeting, we heard from our Nitrogen Cycle working group. In this meeting, we heard from some members of the team about their research on the complex relationships between nitrogen, phosphorus and carbon in various ecological contexts.
Re-examining phosphorus limitations on photosynthesis and productivity in tropical forests- Ning Dong
Ning discussed preliminary results regarding phosphorus limitation in tropical forests, emphasizing their crucial role in tropical ecosystems. There is an ongoing debate regarding the limitations of nitrogen and phosphorus on the productivity in tropical forests, some studies show both N and P are limited (Wright et al., 2018 Ecology) whilst others only have detected phosphorous limitation (Cunha et al. 2022. Nature). The mechanism of driving productivity is still unknown. Effect Traits are traits that determine the effects of plants on ecosystem functions, which are still incorrectly represented in ecosystem models, hampering their predictive skill. We have re-examined with a newly compiled gas exchange dataset that was designed to quantify the dependence of photosynthetic capacities (Vcmax25, Jmax25) on leaf N and P concentrations – from an alternative perspective, considering leaf nutrients as consequences, rather than causes, of leaf function. We found that leaf N per unit area can be expressed as a linear combination of components related to leaf mass per area and Vcmax25, whereas leaf P per unit area can be expressed as a linear function of Jmax25. We also found that there is no soil P limitation on photosynthesis and both climate and soil controls on leaf economic traits at species and community levels. These findings provide a promising route towards an optimality-based approach to modelling leaf traits and their relationships to properties of climate and soils and underlaying drivers of productivity in tropical forest.
The cost of acquiring and using nitrogen relative to water (ß) modifies leaf nitrogen content through two primary pathways- Evan Perkowski, delivered by Nick Smith.
The focus was on investigating the relationship between environmental factors, soil data, and leaf traits in Texas. The state exhibits a precipitation and nutrient gradient from east to west. The study collected leaf samples over 2 years from 23 sites and analysed them for nitrogen content and carbon13 isotopes. The ß term and leaf Ci: Ca were calculated along with climatic data. Soil nitrogen availability and moisture were important variables considered. The study found that ß varied significantly across different environmental gradients and species, with C3 and C4 plants showing distinct beta values. Notably, ß decreased with higher soil nitrogen availability, indicating a reduced carbon cost for acquiring nitrogen.
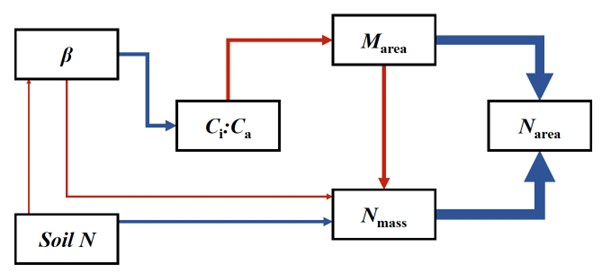
Conversely, ß increased with increased soil moisture due to a reduced carbon cost for acquiring water. Evan developed a structural equation model to explore the drivers of these leaf traits and their relationships. The model revealed a positive relationship between ß and Ci:Ca , influencing leaf mass per area (Marea) which aligns with Wang Han’s model for LMA. This, in turn, affects leaf nitrogen content on an area basis which is also influenced by mass-based leaf nitrogen. Interestingly, there was no direct relationship between Ci:Ca and mass-based leaf nitrogen but ß indirectly decreased mass-based leaf nitrogen. The study suggested that increases in soil nitrogen might be allocated to processes other than photosynthesis. Additionally, the model highlighted the complex pathways (three primary pathways) through which soil nitrogen availability influences leaf nitrogen, two of which are supportive of patterns expected from optimality theory.
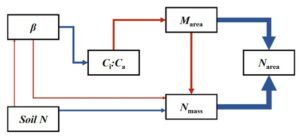
Figure 1. Soil nitrogen indirectly increases leaf N content through three primary pathways.
Empirical constraints and theoretical insights inform a better understanding of ecosystem C and N cycle interactions- Beni Stocker
The focus was on the carbon and nitrogen cycles and their response to environmental changes, particularly climate change and rising CO2 levels. The implementation of coupled carbon and nitrogen cycles in global vegetation models used for the global carbon budget (Trendy v8) was discussed. Beni compared the land carbon sink over the past 60 years between carbon-only models and models with interactive carbon and nitrogen cycles. While the interannual variability was consistent with observations, the carbon-only models showed a narrower spread and better agreement with observations in the mean over the last 10 years. In contrast, the models with interactive carbon and nitrogen cycles displayed divergent trends. There is a need for more observational constraints on the response of the carbon cycle to environmental conditions and how it is affected by nitrogen availability. The traditional modelling approach emphasised a causal relationship between soil nitrogen availability, growth regulation and the plant’s C:N stoichiometry. The presentation proposed a focus on the modification of allocation and the balance between above and below-ground growth, influenced by nitrogen availability and the direct effects of the atmospheric environment (climate and CO2) through the acclimation of photosynthetic efficiency and implications for a plant’s N demand. Developing model formulations and observational benchmarks for predicting and testing plant responses in allocation and acclimation should be a high priority in the development of a next-generation coupled C-N model.
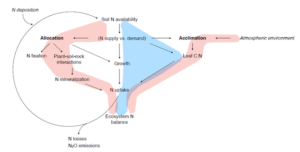
Figure 2. Nitrogen cycling in terrestrial ecosystems
First C-N model results- Beni Stocker
The focus was on the constraints of field observations and ecosystem experiments in formulating models to capture important responses. This study utilised field data, experimental data and published databases of ecosystem manipulation experiments. An overview of response ratios under elevated CO2 and nitrogen fertilisation was given. Under elevated CO2, there was a reduction in Vcmax, and an increase in GPP, NPP and root production. The root-to-shoot ratio also increased, while organic nitrogen uptake decreased. Conversely, under nitrogen fertilisation, there was an increase in NPP, with a larger above-ground increase compared to a below-ground increase, resulting in a decrease in the root-to-shoot ratio. Vcmax shows no change to nitrogen fertilisation. There is a need to predict these responses simultaneously using modelling approaches. The model incorporated functional balance for allocation (optimal root: shoot ratio), acclimation at the leaf level (P-model), N-uptake (based on first principles) and the assumption that canopy-total N in Rubisco scales with absorbed light. The results showed that the model captured the observed responses for elevated CO2 and nitrogen fertilisation.
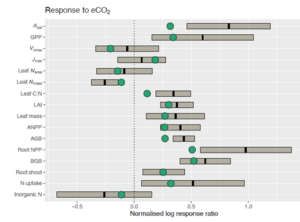

Figure 3. Predicted and observed responses to a) elevated CO2 and b) Nitrogen fertilisation. Stocker et al., (in prep).